

Le Saint Graal des linguistes revisité : le rôle de la génétique dans l’acquisition du langage à l’ère post-chomskyenne
Vasiliki VALLA , Université Aristote de Thessalonique, Grèce
Résumé
Dans cet article, on tentera d’expliquer au lecteur les théories majeures sur l’émergence du langage et les dernières mises à jour sur la base génétique de l’acquisition du langage, en présentant ainsi la perspective interdisciplinaire de la linguistique évolutive. Pour ce faire, on commence par mettre en évidence les questions fondamentales du domaine, suivies d’un aperçu critique des hypothèses courantes sur l’émergence du langage. Nous continuons avec la description de la connaissance actuelle sur les données neuroanatomiques et génétiques impliquées dans l’acquisition et le développement du langage. Enfin, on partage quelques réflexions sur la manière et la mesure dans laquelle les processus de l’évolution culturelle ont influencé l’évolution de la structure linguistique et l’émergence du langage lui-même.
Acquisition du langage - Approche biolinguistique - Approche nativiste - Émergence du langage - Gène FOXP2 - Génétique - Linguistique évolutive
Abstract
In this essay, we try to provide the reader with a connective tissue between the major theories on language emergence and the latest updates on the genetic basis of language acquisition, thus providing proof of the interdisciplinary perspective of evolutionary linguistics. To do so, we begin by highlighting the fundamental questions of the field, followed by a critical overview of the mainstream hypotheses on language emergence. We continue with an outline of the current knowledge on neuroanatomical and genetic data implicated in language acquisition and development. We proceed with some thoughts on how and to what degree the processes of cultural evolution have been a factor affecting the evolution of linguistic structure and the emergence of language itself.
Biolinguistique approach - Evolutionary linguistics - FOXP2 gene - Genetics - language acquisition - language emergence - Nativist approach
1. DÉCOUVERTE DE LA LINGUISTIQUE ÉVOLUTIVE
D’après Descartes (Kenny, 1970 in Corballis, 2017), les animaux étaient des machines fonctionnant selon des principes mécaniques, alors que les êtres humains étaient équipés d’une liberté de pensée capable de défier la mécanique. Ceci était plus évident, pensait-il, dans le langage lui-même. Ainsi, dans une lettre de 1646 adressée au Marquis de Newcastle, il écrit :
Il n’y a aucune de nos actions extérieures, qui puisse assurer ceux qui les examinent, que notre corps n’est pas seulement une machine qui se remue de soi-même, mais qu’il y a aussi en lui une âme qui a des pensées, excepté les paroles […] (1953 : 1254-1257).
Le langage humain est sans doute la caractéristique principale de notre espèce. Nous nous sommes demandés si notre seule espèce était douée du langage, pourquoi le langage était tel qu’il est et comment il a été créé. Des explorations préliminaires sur ces questions remontent à l’ouvrage de Charles Darwin, l’« Origine des Espèces », publié en 1859. Mais la plupart des hypothèses basées sur les conditions scientifiques du XIXe siècle étaient dépourvues de vérifications empiriques ou expérimentales suffisantes. Notons que la Société de Linguistique de Paris a interdit la discussion de ces questions scientifiques en 1866. Après presque un siècle de silence, à partir des années 1950, l’intérêt pour ces questions s’est renouvelé dans les milieux linguistiques, grâce à la constitution de grandes collections de données linguistiques, à la compréhension du comportement des êtres humains et des autres animaux, ainsi qu’à la contribution importante de plusieurs autres disciplines.
La discipline, relativement nouvelle de la linguistique évolutive (Hauser et al., 2007) vise à identifier quand, où et comment le langage prend naissance, change et disparaît (Ke & Holland, 2006) et quelles sont les interférences entre l’acquisition du langage et la diversité langagière (Clark, 2003). Étant donné la difficulté de récupérer des comportements linguistiques à partir d’archives fossiles (Lieberman, 2006), nous pouvons juste supposer à quoi ressemblaient les langues primitives des premiers hominini et de quelles capacités ces hominini disposaient, afin de pouvoir les étudier. Par conséquent, des études en linguistique évolutive, en particulier celles sur l’origine des langages, se sont largement limitées à des données synchroniques. Cependant, la perspective interdisciplinaire du domaine permet, dans une certaine mesure, de surmonter le problème en basant ses découvertes sur des preuves empiriques obtenues par la recherche dans d’autres disciplines.
Ainsi, par exemple, les archives fossiles de nos ancêtres, mises au jour en Afrique du Nord, révèlent que notre espèce a évolué vers sa forme actuelle il y a environ 150 000 ans (White, 2003). Des documents archéologiques constituent des points de référence aux grandes migrations humaines, comme celle qui se sont réalisées par voie maritime (Coupe, 2002) et qui reflètent, de manière indirecte, l’enrichissement et le raffinement des capacités de communication dont les ancêtres disposaient durant ces périodes. Par ailleurs, la combinaison de la génétique moléculaire et de la neuroscience contribue à l’identification de certains développements importants liés au langage dans le génome humain (Fisher & Marcus, 2006, 2010 ; Fitch, 2010 ; Graham & Fisher, 2015 ; Deriziotis & Fisher, 2017).
1.1. Questions fondamentales en linguistique évolutive
La notion du Language Trait chez l’espèce humaine est caractérisée par deux éléments.
Le premier élément est celui que l’on appelle la capacité biologique de langage, qui comprend les fonctions élémentaires de certains organes physiques et des compétences cognitives pour le traitement des matériaux linguistiques (canal vocal-auditif, mémoire associative). Une question majeure est de savoir si cette capacité est spécifique au langage (Chomsky, 1986 ; Pinker & Bloom, 1990) ou si elle provient de capacités générales à travers l’exaltation (Gould & Vrba, 1982). De nombreuses études évaluent la mesure dans laquelle la capacité innée d’apprentissage et d’utilisation d’une langue est propre à l’homme (Lieberman, 2006). Elles sont souvent classées dans les études sur l’évolution du langage.
Le deuxième élément concerne les idiolectes et le langage communautaire. Selon Kirby (2007), les idiolectes sont le corps de la connaissance, attestés dans la parole, que les individus utilisent pour traiter une langue. Le langage communautaire est le consensus des idiolectes, grâce auquel les locuteurs d’une population communiquent entre eux avec un succès habituel (Mufwene, 2008). Une question fréquemment posée est de savoir comment les idiolectes ou les langages communautaires interagissent durant le contact linguistique (Thomason & Kaufman, 1988) ou durant la transmission culturelle (Christiansen & Kirby, 2003).
La linguistique évolutive traite également de l’identification des processus réels qui mutent au cours de l’évolution du langage. Il existe des propriétés invariantes observées dans les langages. Par exemple,
– des significations contenues dans des unités discrètes (sons ou gestes) sont attribuées de façon arbitraire (Hockett, 1960) ;
– parmi les ordres possibles des constituants nucléaires d’une clause transitoire (à savoir le sujet (S), le verbe (V) et l’objet (O)), seuls les SOV, SVO, VSO apparaissent fréquemment dans les langues du monde (Greenberg, 1963) ;
– le langage peut utiliser des matériaux minimaux pour maximiser son pouvoir d’expression à travers la récursivité (Chomsky, 1986 ; Pinker, 1990, 1995 ; Fitch, 2010).
Ces caractéristiques particulières de la structure et de l’usage linguistique qui sont présentes dans la plupart des langues du monde, mais pas nécessairement dans toutes, sont considérées comme des universaux linguistiques (Greenberg, 1963 ; Christiansen & Kirby, 2003). L’étude des universaux et de leurs causes peut nous renseigner sur la manière avec laquelle le langage évolue, sur ses restrictions et sur la façon dont il est relié à d’autres capacités du cerveau humain (Hagoort, 2017 ; Moreno et al., 2018 ; Boris, 2017).
Il est clair que ces questions ainsi que toutes les autres abordées par la linguistique évolutive (Gong, 2014) doivent s’appuyer sur une recherche linguistique solide et c’est pour cette raison que cette dernière doit dépasser ses frontières traditionnelles et mettre à profit les découvertes scientifiques d’autres domaines afin d’établir des arguments convaincants sur les questions fondamentales du domaine. Dans une publication récente, Berwick & Chomsky (2017) déclarent :
Languages change, but they do not evolve. It is unhelpful to suggest that languages have evolved by biological and non biological evolution – James Hurford’s term. The latter is not evolution at all.
Ce point de vue est assez commun en linguistique, mais un grand nombre de linguistes évolutifs contestent le fait qu’une perspective évolutive soit non seulement appropriée mais aussi notre meilleure option pour comprendre l’évolution du langage (Croft, 2000 ; Mufwene, 2008 ; Steels, 2011 ; Hurford, 2012). Le tableau 1 figure cette sorte de parallélisme entre l’évolution biologique et l’évolution du langage.
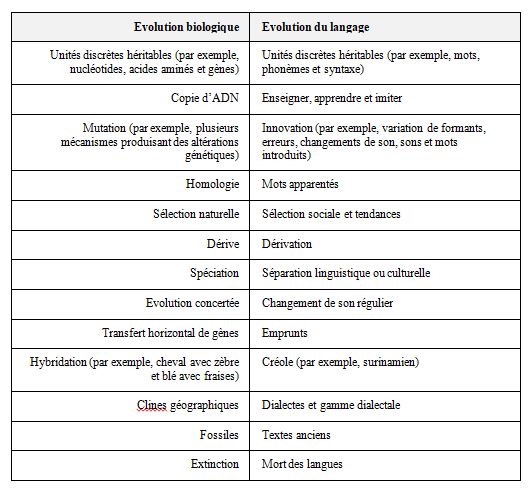
Tableau 1 – Similitudes entre évolution biologique
et évolution du langage (adapté de Pagel, 2009)2. DEUX POINTS DE VUE OPPOSÉS SUR L’ ÉMERGENCE DU LANGAGE
Bien que les origines et la complexité du langage aient été discutées avant l’entrée de Chomsky dans le monde de la linguistique, ce n’est qu’après sa réponse à Skinner (Chomsky, 1959) que les processus impliqués dans l’acquisition du langage ont fait l’objet de débats linguistiques intenses. Le mystère de l’acquisition du langage qui a préoccupé des psychologues, des linguistes et même des philosophes (Skinner, 1957, Chomsky, 1959) concernait le fait que le langage semble être réservé uniquement au genre humain. Les points de vue divergents sur l’acquisition du langage entre Chomsky et Skinner se sont maintenus jusqu’à aujourd’hui et concernent les mécanismes par lesquels les êtres humains acquièrent le langage. Le problème central de leur désaccord se manifeste dans l’importance accordée à l’environnement linguistique et à la contribution au langage chez l’enfant. Dans cet article, on souligne les traits principaux de deux grandes théories de l’acquisition du langage : la théorie nativiste et la théorie empiriste, fondée sur l’usage. Selon la première théorie, la compétence grammaticale est innée : une capacité qui est spécifique à l’être humain. L’approche alternative de l’acquisition du langage met, quant à elle, l’accent sur les processus cognitifs généraux des enfants et considère le processus d’acquisition comme un développement progressif.
2.1. L’approche nativiste de l’acquisition du langage
La discussion sur les mécanismes liés à l’acquisition du langage remonte à la réponse de Chomsky à l’idée de Skinner selon laquelle le comportement verbal des enfants est prévisible parce qu’il est acquis par l’intermédiaire de stimuli externes, renforçateurs (Chomsky, 1959). À l’époque, la réponse de Chomsky a pu être considérée comme par trop radicale, et pourtant aujourd’hui, l’argument selon lequel il existe quelque chose d’inné dans notre capacité d’acquérir et de traiter le langage, est considéré comme valide dans les milieux linguistiques. Une des principaux arguments est que le langage humain est trop complexe pour que la seule perception de l’environnement et l’exposition à la langue puissent expliquer une acquisition aussi rapide par l’enfant. Par conséquent, la grammaire, en tant qu’ensemble complexe de règles gouvernant l’usage du langage, doit être innée. Une idée était que toutes les langues partagent des similitudes sous-jacentes fondamentales telles que la présence de catégories grammaticales, comme les noms et les verbes, ce qui serait sûrement expliqué par la prédétermination génétique. La seule façon de remettre en question cette idée d’innéité est d’offrir une « general learning strategy » qui pourrait justifier l’acquisition, mais qui, selon Chomsky (2006), n’a pas encore été proposée. La grammaire universelle apparaît comme une théorie qui explique ces « universaux linguistiques » (Cook & Newson, 2007).
Cependant, la notion de grammaire universelle n’est pas acceptée par toute la communauté linguistique (Evans, 2009 ; Everett, 2005 ; Dunn, 2011 ; Müller, 2013). Evans et Levinson (2009) présentent plusieurs aspects dans lesquels les langues diffèrent fondamentalement plutôt que superficiellement et l’une des différences entre les langues est l’ordre des mots. Certaines langues ont un ordre SVO, d’autres un ordre SOV et il peut même y avoir des changements importants dans une même langue à travers le temps. Selon Pinker (1995), l’anglais, par exemple,
Has changed from a free-word order, highly inflected, topic-prominent language, as its sister German remains to this day, to a fixed-word-order, poorly inflected, subject prominent language, all in less than a millennium
Comme attendu, les langues diffèrent également au niveau de la phonologie, de la morphologie et de la sémantique. La plupart de ces différences sont souvent considérées comme superficielles par les partisans de la grammaire universelle. Néanmoins, la variation grammaticale parmi les langues s’étend au-delà de l’ordre des mots. Ainsi, par exemple, plusieurs langues ne disposent pas de classes d’adverbes ouvertes (Evans, 2009). De plus, certaines restrictions trouvées en anglais ne s’appliquent pas à d’autres langues telles que l’italien ou le russe, ce qui réfute non seulement l’existence des universaux linguistiques, mais réfute également la position anglo-centrique que Chomsky adopte (Evans, 2009).
Si les affirmations nativistes étaient poussées à l’extrême, on pourrait dire que les enfants seraient linguistiquement compétents dès les premiers stades de leur vie, mais le manque de performance est dû évidemment à des compétences insuffisamment développées, liées à la motricité, à la pratique, aux ressources insuffisantes et à d’autres aspects performatifs de l’usage du langage. Cependant, l’hypothèse d’une compétence linguistique complète dès la naissance remet en question le rôle fonctionnel que la cognition générale aurait dû avoir dans l’acquisition du langage. Selon le paradigme nativiste, « l’input » n’est pertinent que dans le sens où il aide l’enfant à appliquer les catégories et règles grammaticales innées au Dispositif d’acquisition du langage (LAD).
Au sein de la faculté du langage, deux composantes majeures ont été définies : la Faculty of language-broad sense (FLB), qui comprend les systèmes conceptuel-intentionnel et sensori-moteur et la Faculty of language-narrow sense (FLN) (Hauser, 2002).Ces deux systèmes constituent certaines parties de la cognition générale, mais FLB inclut FLN, qui est indéniablement l’élément controversé dans le schéma. De plus, ces deux systèmes internes sont attribués à la seule race humaine. Cette affirmation a reçu beaucoup de critiques, car elle soutient effectivement que la faculté du langage est complètement autonome et n’a aucun lien avec la cognition ou l’intelligence. Néanmoins, la théorie nativiste est plutôt claire sur ces points aujourd’hui et la partie liée à l’intelligence a été enlevée de l’équation de l’acquisition. En établissant une analogie entre les animaux et les imbéciles qui acquièrent le langage, Chomsky (2006) affirme en effet que l’intelligence est un mécanisme séparé. L’argumentation mise en avant est celle du « device » spécial, duquel l’évolution est ressortie, et qui nous dote du langage et de la grammaire dès la naissance.
L’argument en faveur d’une faculté du langage biologiquement dotée, est également affaibli par les affirmations d’Everett (2005), qui dit que la langue amazonienne Pirahã ne permet pas la récursivité, une des caractéristiques fondamentales du langage selon la théorie de Hauser (2002). Selon la réponse de Hauser (Hauser, 2002), aucune autre espèce que l’homme n’a cette capacité de recombiner des unités significatives en une variété illimitée de structures plus grandes, chacune différant systématiquement au plan du sens. Certes, les systèmes communicatifs peuvent être retrouvés chez diverses espèces animales, mais quelque chose d’aussi complexe que le langage est resté inobservable en dehors de l’espèce humaine. Par conséquent, souligner l’absence de récursivité dans les systèmes communicatifs des animaux est pleinement justifié et même totalement inutile. Pinker (2005) a critiqué la plupart des arguments fournis par Hauser en 2002, en disant que la récursivité peut être observée ou interprétée dans d’autres fonctions sensorielles chez l’homme, tels que la cognition visuelle : le monde extérieur est perçu comme étant constitué d’éléments discrets, qui peuvent être réunis pour former des constituants plus grands, et les séquences qui sont observées comme des paires ou des groupes peuvent être sans fin. Il est toujours possible de créer des constituants plus grands à partir de divers éléments. Puisque l’univers lui-même est infini et que les humains ont tendance à catégoriser, il est probable que ce que l’on appelle « discrete infinity » soit perceptible partout. Si nous devions intégrer dignement la propriété de la récursivité dans l’approche nativiste, nous devrions attribuer la propriété à la FLB plutôt qu’à la FLN.
2.2. L’approche basée sur l’usage
L’approche basée sur l’usage, telle que soutenue par Tomasello, met l’accent sur l’opposition nurture vs nature (en francais : le débat inné-acquis) (Tomasello 2000, 2003, 2008). C’est la cognition générale de l’enfant qui est considérée dans cette approche, plus que la capacité proprement linguistique. Selon Tomasello, les enfants n’atteignent pas la phase d’acquisition du langage avant l’âge d’un an. Jusqu’à ce moment-là, ils développent des compétences telles que l’intention-reading (dimension fonctionnelle) et pattern finding (dimension grammaticale), qui sont toutes les deux des fonctions cognitives généralement sans rapport avec le langage. Tomasello considère l’intention-reading comme spécifique à l’espèce, ce que les nativistes trouvent extrêmement important. Cependant, on pourrait dire que l’intention-reading est inobservable chez les primates car elle implique la lecture des intentions des autres et il semble que l’action intentionnelle soit uniquement réservée aux espèces capables d’avoir l’intention de faire quelque chose, ce qui signifie qu’elle est cognitivement avancée. En supposant que l’action intentionnelle existe en effet chez d’autres primates ou animaux, il semble peu probable que le reste du règne animal réponde aux exigences cognitives pour prendre en compte les objectifs des autres ou pour comprendre que les autres agissent intentionnellement. L’acquisition de tout élément linguistique dépend d’une telle compréhension.
L’écart entre la compréhension et l’acquisition, ce que Chomsky appelle chasm, et le comblement de cet écart, sont les points de divergence fondamentale entre les deux théories. D’après Tomasello (2008)
Almost all infants communicate by pointing before they have acquired any productive language, and many also use some kind of iconic or conventionalized gestures as well […] This suggests that human pointing and other gestures may already embody forms of social cognition and communicative motivation that are unique to the species, and that are necessary as a first step on the way to linguistic conventions both phylogentically and ontogenetically
À l’instar de Chomsky, Tomasello reconnaît que certains mécanismes sont uniques chez l’homme. Néanmoins, ces mécanismes sont plutôt le sous-produit d’un système cognitif plus évolué chez l’homme. Naturellement, la cognition ou l’intelligence pourraient être considérées comme innées dans une certaine mesure, mais il vaut mieux éviter le terme lui-même en raison de ses implications spécifiques à l’espèce. Tandis que l’intention-reading est considérée par Tomasello comme caractéristique unique d’un être humain, il s’agit plutôt d’un effet secondaire d’une cognition générale plus avancée, comme indiqué précédemment. Cette théorie accorde de l’importance à la communication pré-linguistique, sur laquelle la communication linguistique est ensuite construite. Afin de comprendre comment fonctionne l’acquisition du langage, il faut comprendre la façon dont fonctionne la grammaire. Il semblait que la première chose que l’enfant apprend dans une langue est le mot ; mais d’après cette théorie, c’est tout le contraire. Alors que le mot est ce qui est articulé en tout premier, ce n’est certainement pas le premier qui est appris. L’enfant écoute d’abord les conversations, les énoncés et à l’aide de l’observation et de la déduction, l’enfant essaie de comprendre le sens d’un mot. C’est la fonction du mot dans une phrase, ainsi que la signification des éléments environnants, qui guideront la tentative analytique de l’enfant pour découvrir le sens du mot. Il est clair qu’on ne peut pas comprendre le sens d’un mot juste en analysant un seul énoncé. L’enfant se souvient de la série des énoncés dans lesquels le mot particulier a été mentionné et, comme en mathématiques, l’enfant découvre le sens de la constante (le mot) à l’aide de plusieurs équations (énoncés).
D’après Tomasello, la deuxième compétence cognitive d’une importance considérable à l’acquisition du langage est le pattern-finding. La première différence entre pattern-finding et intention-reading est que le pattern-finding est répandu chez tous les primates puisqu’il représente/décrit le processus analytique qui consiste à trouver des régularités et à extraire les données pertinentes de l’input, c’est-à-dire du langage. La récursivité semble être considérée comme un sous-produit des restrictions cognitives de l’être humain ainsi que des limitations pragmatiques liées au discours. Avant d’accepter l’universalité de la grammaire, il faut d’abord prendre en considération le fait que les compétences cognitives et la même pensée analytique sont également universelles, ce qui permet de considérer l’universalité de la grammaire comme un sous-produit des processus énoncés.
3. ASPECTS ANATOMIQUES ET GÉNÉTIQUES DE L’ÉVOLUTION DU LANGAGE
L’anatomie est liée à l’évolution du langage car cette capacité propre à l’homme dépend notamment de l’importance de la taille et du degré de complexité de la structure de son cerveau. Des études classiques ont trouvé que le cortex comprend deux zones dédiées au langage, ce qui est prouvé par la diminution de la fonction du langage quand elles ont été endommagées (Fisher, 2006). Des études de neuro-imagerie montrent que d’autres zones corticales sont également impliquées dans le langage (Fedorenko, 2009 ; Armeni, 2017 ; Hagoort, 2017 ; Mars, 2018), mais on mettra l’accent sur les deux régions classiques car elles déterminent la morphologie du crâne. La parole et, par conséquent le langage, dépendant d’une série des traits anatomiques (par exemple, la taille du canal hypoglosse sur la base de l’os occipital, la forme de l’os hyoïde (langue), le diamètre des vertèbres etc.) peut être étudié à travers les découvertes paléontologiques (fossiles). Klein (2017) procède à un examen approfondi des phases successives, anatomiques et archéologiques, de l’évolution humaine en faisant la liaison avec l’évolution du langage. Selon ses conclusions, l’anatomie de la parole était probablement en place, au moins sous une forme rudimentaire, il y a 2,5 à 2Ma [1], mais le langage entièrement développé est probablement apparu il y a environ 50.000 ans. Comme le remarque Klein, l’élaboration lente mais progressive des éléments archéologiques de 2,5 à 50 ka [2] peut signifier le développement parallèle de formes de langage beaucoup plus sophistiquées mais encore primitives.
Les preuves apportées par des fossiles à partir des moulages crâniens et de la structure osseuse du canal vocal, de la colonne vertébrale thoracique et du système auditif, suggèrent qu’il y a eu une phase évolutive passant d’une condition simiesque chez les australopithèques, qui ont vécu essentiellement il y a 2Ma, à une condition essentiellement humaine chez les membres du genre Homo, qui ont vécu essentiellement ensuite (Everett, 2017). Pour des raisons anatomiques, les australopithèques n’étaient probablement pas capables de parler aussi bien que les chimpanzés, et ce manque probable de parole pourrait être ajouté à d’autres aspects simiesques de leur anatomie et de leur comportement pour permettre de conclure qu’il s’agissait essentiellement des singes bipèdes (bipedal apes). D’un autre côté, la possibilité de parole pourrait être ajoutée à d’autres traits qui suggèrent que toutes les espèces d’Homo, après2 Ma, pourraient raisonnablement être appelées humaines.
L’émergence d’Homo il y a 2,5-2Ma est encore mal documentée, mais elle a coïncidé avec la plus ancienne indication de la dépendance des Homini à la technologie de la pierre taillée. Même les plus anciens outils de pierre taillée incontestables, attribués à l’Oldowayen entre environ 2,6 et 1,7 Ma, exigeaient que les fabricants maîtrisent les mécanismes de la pierre taillée (Stout, 2009 & 2012). Les principes ne sont pas intuitifs et ne peuvent être maîtrisés par les chimpanzés. Des observations expérimentales confirment que les humains vivants peuvent apprendre des techniques de taille simples par imitation, mais ils apprennent de manière beaucoup plus efficace et fiable s’ils reçoivent une instruction verbale (Everett, 2017 ; Stout, 2012 ; Bar-Yosef, 2017). Le langage rudimentaire est donc susceptible d’expliquer à la fois la facilité avec laquelle les gens de l’Oldowayen semblent obtenir des éclats à partir d’un même noyau, et leur capacité à transmettre fidèlement leur connaissance de génération en génération pendant des centaines de milliers d’années.
La recherche devient encore plus compliquée si on prend en considération que des formes de langage encore primitives étaient probablement liées à l’élaboration progressive post-Oldowayenne de la technologie de la pierre taillée, débutant il y a environ 1,76 Ma avec les premiers bifaces et d’autres outils façonnés de la Tradition Acheuléenne, suivies de bifaces plus raffinées de la tradition acheuléenne, et puis, il y a 300-250 ka, de la disparition des bifaces et de leur remplacement au Middle Stone Age en Afrique et au Paléolithique moyen à l’ouest de l’Eurasie par des outils de pierre taillée retouchés (bords modifiés) (Stout, 2011). Des fragments occasionnels d’ocres, des coquilles d’escargots intertidales perforées et des fragments incisés de coquille d’œuf d’autruche peuvent signifier que le « symbolisme », intimement lié au langage historiquement, est apparu à la fin du Middle Stone Age, il y a 100-60 ka, mais si les preuves archéologiques sont acceptées, cela signifie une forme de symbolisme qui était qualitativement différente du genre univoque qu’on connait historiquement. Le genre historique, marqué par l’art irréfutable et les ornements personnels, est apparu il y a 50-40 ka, ce qui semble indiquer que c’était aussi la date de l’apparition du langage développé.
L’apparition abrupte du langage et d’autres traits cognitifs complètement modernes il y a 50-40 ka s’est faite sûrement en Afrique, et une cognition accrue est susceptible d’expliquer l’expansion presque simultanée des Africains complètement modernes en Eurasie, qui ont remplacé ou submergé les Néandertaliens et autres humains non modernes (Everett, 2017 ; Tatersall, 2017). L’ADN ancien pourrait confirmer cette hypothèse, s’il était possible de déterminer si les Néandertaliens et d’autres personnes non modernes manquaient de gènes qui étaient fondamentaux pour le langage et d’autres fonctions cognitives chez les humains historiques.
4. GÈNES ET ÉVOLUTION DU LANGAGE
Alors que la plupart des animaux utilisent la communication vocale, les êtres humains sont uniques dans l’utilisation de la langue écrite. Cela implique le développement d’un système de décodage parmi les sons, les symboles et les concepts. Comme le suggère Mattingly (1972), « reading is parasitic on speech » car la lecture dépend de toutes les composantes du langage parlé : la syntaxe, la morphologie, la phonologie, la pragmatique et le lexique (van der Lely, 2014 ; Hulme, 2013). La relation étroite entre les compétences linguistiques à l’écrit et à l’oral est bien acceptée et particulièrement évidente dans la comorbidité dans les troubles du langage et la dyslexie développementale (DD) (Jiménez-Bravo, 2017 ; Peterson, 2012). Les troubles du langage et la dyslexie développementale sont des troubles neurodéveloppementaux communs, caractérisés par des difficultés inattendues liées au langage verbal et à la lecture, respectivement, malgré les opportunités éducatives et socioéconomiques adéquates, ainsi que d’autres méthodes de développement normal (Plaza, 2016).
Au cours des dernières années, des études de génétique moléculaire dans des contextes familiaux ou cas-témoins ont identifié des gènes candidats pour les troubles du langage et la dyslexie développementale, avec plusieurs facteurs de risque génétiques contribuant à ces deux conditions (Mozzi, 2016 ; Carrion-Castillo, 2013 ; Paracchini, 2011). Le premier gène à être impliqué dans un trouble sévère lié à la parole et au langage est le FOXP2, qui semble avoir muté dans une grande famille atteinte de dyspraxie verbale (Lai, 2001). Une fois identifié, le rôle du FOXP2 dans les capacités (troubles) linguistiques a été confirmé de manière indépendante dans plusieurs études (Graham, 2013, Fisher, 2009 ; Derviziotis, 2017), et une analyse évolutive de sa séquence codante a révélé deux substitutions d’acides aminés spécifiques à l’homme. Cela a conduit à l’hypothèse que des changements récents dans la protéine du FOXP2 ont contribué au développement des compétences verbales chez l’homme (Enard, 2002). Cette possibilité a été confirmée par des études sur des modèles animaux et lignées cellulaires (Enard, 2011, Konopka, 2009) mais contestées par d’autres observations (Derviziotis, 2017 ; Janik, 2014 ; Krause, 2007).
La découverte que les Néandertaliens avaient déjà les variantes du FOXP2 spécifiques à l’homme (Krause, 2007) a alimenté des hypothèses sur leur impact (ou leur absence) sur le développement du langage et sur le timing de l’origine du langage moderne (Ackermann, 2014 ; Johansson, 2014). En fait, il existe encore un débat considérable concernant l’existence d’un système de communication utilisé par les humains archaïques, comparable à celui des humains modernes (Klein, 2017 ; Ackermann, 2014 ; Johansson, 2014 ; Dediu, 2013 ; Berwick, 2013a & 2013b).
Dix gènes associés à un trouble du langage ou à une dyslexie développementale ont été étudiés pour leurs implications évolutives potentielles (Mozzi, 2016). L’étude consiste à comparer les homologues de gènes entre H. sapiens et d’autres vertébrés, et maintenant entre H. sapiens et les hominines eurasiennes disparues, H. neanderthalensis et les « Denisovans », pour lesquels l’ADN est disponible. H. sapiens, venu d’Afrique, a remplacé les deux espèces il y a environ 50 ka. Contrairement aux Néandertaliens dont l’anatomie est bien connue à partir des fossiles, les Denisovans sont presque entièrement connus à partir de l’ADN, tiré de quatre fossiles morphologiquement non diagnostiqués, trouvés dans la grotte de Denisova, au sud de la Sibérie (Meyer, 2012 ; Reich, 2010). Les Denisovans n’ont pas de place officielle dans le système linnéen, mais ils peuvent être représentés par des fossiles assignés à H. Erectus en Chine. Ces fossiles remontent à environ 300 et 50 ka (Klein, 2017). Selon l’ADN, les Denisovans et H. neanderthalensis étaient des espèces sœurs qui partageaient un ancêtre commun avec H. sapiens il y a environ 700 ka.
4.1 Gène FOXP2
Le premier gène à être impliqué dans un trouble sévère lié à la parole et au langage était donc le FOXP2. Une fois découvert, le facteur de transcription FOXP2 (Fig.1) a été un paradigme pour l’association des gènes, des neurones, du cerveau et du langage parlé (Graham, 2013, Fisher, 2009 ; Derviziotis, 2017). Des approches moléculaires dans les systèmes cellulaires ont montré la régulation des gènes (Enard, 2011 ; Konopka, 2009 ; Krause ; 2007) et l’interaction avec des protéines (Estruch, 2016 ; Vernes, 2008 ; Li, 2004) impliquant le FOXP2 dans les processus neuronaux tels que la différenciation, l’excroissance des neurites, le guidage axonal et la plasticité synaptique. Curieusement, un sous-ensemble de ses cibles et/ou partenaires d’interaction a été associé à d’autres phénotypes neurodéveloppementaux, tels que le SLI (Vernes, 2008), l’épilepsie (Roll, 2010 ; Strauss, 2006), la schizophrénie (Walker, 2012) et les ID/ASD (Deriziotis, 2017 ; Sollis, 2016 ; Chen, 2016 ; Lozano, 2015 ; Lefevre, 2013 ; Mukamel, 2011). Le langage est uniquement humain, mais le FOXP2 a une histoire profondément évolutive et est similaire, parmi divers vertébrés, à une expression conservée, particulièrement dans les sous-types neuronaux dans le cortex, le thalamus, les ganglions de la base et le cervelet (Fisher, 2006). Le Tableau 3 résume l’expression du FOXP2 dans le cerveau. Ainsi, les modèles animaux offrent des indices sur les rôles neuronaux du FOXP2 (French, 2014 ; Wohlgemuth, 2014). Par exemple, des souris avec des mutations correspondant à celles qui causent un trouble de la parole chez l’homme ont montré un trouble d’apprentissage de la motricité (Groszer, 2008) et un séquençage altéré des vocalisations ultrasoniques (Chabout, 2016). De plus, des études électrophysiologiques de ces souris mutantes ont trouvé des effets sur la décharge et la plasticité des circuits neuronaux où le gène est exprimé (French, 2012 ; Groszer, 2008). D’autres indices proviennent d’essais menés sur le diamant mandarin, une espèce aviaire dans laquelle les mâles apprennent à chanter une chanson particulière d’un oiseau tuteur adulte (Wohlgemuth, 2014). Lorsque l’interférence ARN a été utilisée pour supprimer l’expression de l’orthologue du diamant mandarin du FOXP2 dans la zone X, un ganglion de la base impliqué dans l’apprentissage de la chanson, cela a perturbé la modulation développementale et sociale de la variabilité de la chanson (Wohlgemuth, 2014 ; Murugan, 2013). Des études chez d’autres espèces proposent que les contributions de ce gène à la parole et au langage humain sont construites sur des fonctions évolutivement anciennes dans la modulation de la plasticité d’un sous-ensemble de circuits cérébraux (Fisher, 2006 & 2010). Les rôles du FOXP2 sont complexes et variés, même au sein du système nerveux, et il reste encore beaucoup à apprendre sur ses fonctions fondamentales et sur la raison des effets disproportionnés des perturbations sur les capacités linguistiques. Néanmoins, ceci montre comment un gène impliqué dans une forme rare de trouble peut fournir des informations sur les voies neurobiologiques pertinentes.
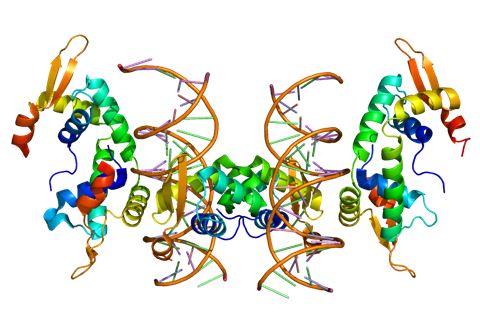
Fig. 1 – Structure de la protéine FOXP2 liée à l'ADN (Base EBI-PDBsum [[Pictorial database of 3D structures in the Protein Data Bank (Base de données picturale de structures 3D dans la banque de données sur les protéines de l’Institut européen de bioinformatique (EBI)), http://www.ebi.ac.uk/thornton-srv/databases/cgibin/pdbsum/GetPage.pl?pdbcode=index.html]]).

Tableau 2 – Expression du FOXP2 dans le cerveau
En ce qui concerne les autres gènes du langage (Mozzi, 2016), le KIAA0319 diffère entre les hominidés et les autres singes, mais pas entre les humains contemporains d’un côté et les Néandertaliens et les Denisoviens de l’autre côté. Comme le FOXP2, le ROBO1, le ROBO2, le SRPX2, le TSC1 et le CNTNAP2 ne diffèrent entre les contemporains et les Néandertaliens que dans les régions non codantes, et la mutation qui a produit la forme moderne, pourrait ainsi améliorer la capacité du langage. Quand les fonctions des gènes du langage connus seront mieux comprises, on pourra apporter plus de lumière sur l’évolution du langage révélée par la plupart des aspects de l’anatomie et l’émergence de nouvelles découvertes pourra nous indiquer quand les langues véritablement modernes sont apparues.
4.2 Neuro-imagerie et génomique : avenir de la linguistique évolutive ou chimère ?
Face à de petites tailles d’effets lors de l’association de variantes génétiques communes avec des mesures comportementales, les chercheurs ont cherché à se rapprocher de la biologie en étudiant les endophénotypes cérébraux. Le domaine de la génomique qui évolue rapidement cherche des corrélations entre les variations interindividuelles dans les domaines de la structure et/ou de la fonction cérébrale – indexées par la neuro-imagerie – et la variabilité au niveau moléculaire – indexée par le génotypage – (Bearden, 2017). Au début on espérait que les polymorphismes génétiques auraient des effets plus importants sur la structure et la fonction du cerveau que sur les sorties comportementales/cognitives distales, en négligeant certains problèmes de robustesse rencontrés par les études d’association génétique du comportement traditionnel.
Cependant, des méta-analyses des volumes sous-corticaux chez plus de 30 000 individus montrent que des mesures structurales dérivées de la neuro-imagerie peuvent montrer autant de complexité dans les fondements génétiques que dans les (endo)phénotypes comportementaux, avec des tailles d’effet lors des associations génétiques individuelles qui sont d’une ampleur similaire (Hibar, 2015). Ainsi, les méthodes de neuro-imagerie génétique sont soumises à des contraintes similaires à celles des études d’association classiques, par rapport à la conception de l’étude, à la taille de l’échantillon, aux problèmes de comparaisons multiples, etc. De plus, la complexité croissante et la richesse des données de neuro-imagerie individuelles, par rapport aux données de tests psychométriques par exemple, entraînent une sensibilité plus grande aux faux positifs, surtout si le travail exploratoire est effectué sur de petits échantillons sans ajustements statistiques suffisants (Button, 2013). Les analyses du FOXP2 donnent un avertissement. L’application de la neuro-imagerie à des personnes ayant des troubles liés au langage et à la parole, à cause de rares mutations codantes du FOXP2de grande pénétrance, a révélé des anomalies structurelles subtiles mais importantes, affectant des régions telles que le gyrus frontal inférieur, le striatum et le cervelet (Morgan, 2016). Par la suite, un certain nombre d’études à petite échelle ont proposé que les polymorphismes introniques communs du FOXP2 (tous d’une importance fonctionnelle inconnue) sont associés aux variations de la structure cérébrale. Mais la plus grande étude à ce jour, réalisée sur une population de 1300 personnes indifférenciées, n’a trouvé aucune preuve pour confirmer cette affirmation (Hoogman, 2014). Néanmoins, comme la taille des échantillons continue d’augmenter pour les cohortes avec des mesures robustes de la structure/fonction cérébrale et des données génotypiques correspondantes (Bearden, 2017) la neuro-imagerie génétique pourrait en effet aider à révéler l’architecture génétique en soulignant la variabilité des circuits liés au langage. Par exemple, on doit anticiper les résultats des études GWAS [3] de variabilité individuelle d’épaisseur, de superficie et de volumes de régions corticales importantes impliquées dans la procédure du langage, ainsi que des faisceaux de la substance blanche qui les relient, et de leurs propriétés fonctionnelles. Il sera particulièrement intéressant d’intégrer les résultats de ces études à ceux de GWAS qui concerne les endophénotypes comportementaux liés au (Graham, 2015). Jusqu’à présent, les synergies ont été limitées à l’évaluation de signaux de GWAS spécifiques au langage/lecture pour des effets sur la structure cérébrale (Gialluisi, 2017).
Il est clair que l’architecture génétique qui façonne la maîtrise du langage chez l’individu et dans l’ensemble de la population est hétérogène, impliquant à la fois des variations communes et rares. Pour découvrir de nouveaux gènes liés au langage et pour mieux comprendre la base moléculaire des compétences pertinentes, il faut allier les méthodes de technologie récente de caractérisation génomique à l’intégration de données provenant d’approches complémentaires dans de multiples disciplines, des molécules à la clinique.
À ce jour, les études GWAS sur les phénotypes de la parole, du langage et de la lecture, que ce soit pour des troubles ou des variations normales, ont révélé peu d’associations significatives, et aucune n’a encore été prouvée de façon convaincante. Ce succès limité peut largement refléter un manque de puissance, à cause de tailles d’échantillon insuffisantes pour détecter les polymorphismes d’un seul nucléotide (SNP) avec de petits effets anticipés, dans un contexte de tests multiples massifs à travers le génome. D’autres recherches similaires sur d’autres phénotypes liés au cerveau, tels que la schizophrénie, montrent que certains participants génotypés/phénotypés de l’ordre de dizaines de milliers, seront nécessaires pour des associations robustes (Schizophrenia Working Group, 2014). Une façon d’atteindre l’échantillon nécessaire est de cibler des cohortes indépendantes avec des informations génotypiques existantes sur le génome et pour lesquelles on a déjà collecté des données sur les mesures de la parole, du langage et/ou de la lecture, puis d’essayer d’harmoniser les phénotypes disponibles entre les cohortes pour des méta-analyses ou même pour des méga-analyses.
La seule chose que l’on pourrait conclure est que lorsqu’on pense à la liaison entre la linguistique évolutive et la génomique, il faut toujours garder à l’esprit que le langage n’est pas une dimension unitaire mais un phénotype complexe à facettes multiples, en prenant en considération des aspects qui varient de la richesse lexicale et de la maîtrise de la parole à la compréhension de la grammaire et aux inférences pragmatiques. La figure 2 illustre la complexité anatomique et les interactions fonctionnelles des réseaux cérébraux, qui nous permet de parler et par conséquent de communiquer et d’interagir. Comment ces aspects sont-ils liés les uns aux autres chez l’individu, comment varient-ils entre les individus, et comment les profils changent-ils à différents points de développement ? Ce sont là des questions auxquelles peu de réponses ont été apportées. Les propriétés distinctes des langues parlées dans différentes parties du monde imposent leurs propres limites à la caractérisation phénotypique.
Fig. 2 – Lobes cérébraux
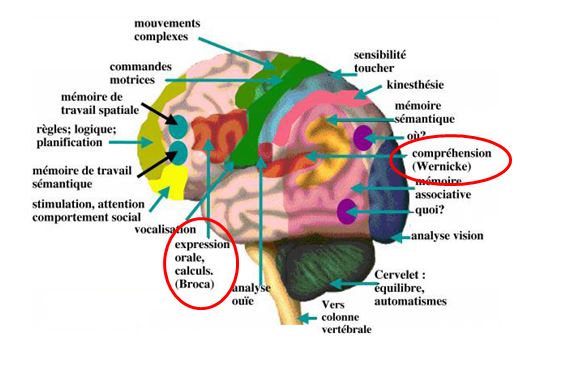
Fig. 3 – Anatomie fonctionnelle du cerveau. Schéma des circuits neuronaux impliqués dans la communication vocale. Les circuits neuronaux impliqués dans l'apprentissage et la production précise de vocalisations comprennent de nombreux réseaux interconnectés dans le cortex, les ganglions de la base et le cervelet. Bien que l'on disposede peu d’informations sur les réseaux de gènes impliqués dans la communication vocale, les dépistages génétiques des individus ayant des troubles de la parole héréditaires et l'examen biologique continu des circuits neuronaux chez les espèces qui utilisent l’apprentissage vocal, comme les passeri, offrent un aperçu du cerveau et des mécanismes génétiques de la communication vocale.
Adapté du site : http://cervenad.blogspot.gr/2014/11/le-cerveau-humain.html5. L’IMPACT CULTUREL SUR L’ACQUISITION DU LANGAGE
Dans un article récent, Fuentes (2016) défend une synthèse évolutive (extended evolutionary synthesis, EES). Il dit qu’une image complète de notre espèce engage le biologique, le culturel et le psychologique simultanément dans le cadre de cette compréhension basée sur l’EES de la condition humaine. En même temps, Everett (2016) aborde la nature de la culture et propose un ensemble de composantes fondamentales de la culture et comment cette notion de culture interagit ensuite avec notre esprit, notre corps et nos relations sociales. Cette conceptualisation de la culture joue un rôle quand on considère que les pressions culturelles auraient pu altérer le développement biologique de l’homme et que ce développement biologique aurait pu à son tour modifier nos cultures, en particulier la façon dont les deux ont contribué à l’évolution du langage (Dediu, 2013 ; Luuk, 2014). Everett (2017) va plus loin en affirmant que le langage (en tant que trialité de chaîne de symboles et de gestes) a évolué au fil d’une progression sémiotique et que cette progression a commencé par des figurations et s’est terminée par des grammaires linéaires non récursives, non hiérarchiques (les grammaires hiérarchiques et récursives sont des embellissements secondaires du langage et ne sont ni nécessaires, ni suffisantes pour le langage). L’auteur dit que le langage a probablement commencé avec H. erectus et rejette l’idée selon laquelle le langage dépend largement d’une sorte de grammaire. Malgré les affirmations contraires (Berwick & Chomsky, 2017), l’idée que le langage est avant tout un outil culturel pour la construction communautaire (Everett, 2012) devient de plus en plus populaire. Selon Everett, les propriétés computationnelles, telles que la dépendance de structure récursive, ou même la linéarité simple, sont secondaires. Il rejette la conception de Chomsky pour qui
Communication, a particular use of externalized language, cannot be the primary function of language, a defining property of the language faculty, suggesting that a traditional conception of language as an instrument of thought might be more appropriate. At a minimum, then, each language incorporates via its syntax computational procedures satisfying this basic property … We take the property of structure dependence of grammatical rules to becentral.
affirmant qu’il ne prend pas en compte de l’information évolutive, du rôle des gestes et de la culture, de la mise en forme architectonique de la grammaire, enfin de la redondance et de l’ambiguïté en tant que caractéristiques naturelles du langage (Everett, 2012 ; Everett, 2017 ; Piantadosi, 2012).
Everett (2017) considère le langage comme lien de sens (sémantique), conditions d’usage (pragmatique), propriétés physiques d’un inventaire de sons (phonétique), grammaire – syntaxe (structure de phrases), phonologie (structure sonore), morphologie (structure des mots) –, discours et principes organisationnels conversationnels. L’accomplissement ultime du langage est de construire des relations, des cultures et des sociétés et la grammaire contribue considérablement à cette tâche. Mais malgré la perception de Chomsky sur le langage, la grammaire elle-même tend en fait à ne pas être plus importante que les autres composantes du langage mentionnées ci-dessus. En général, Everett (2017) rejette l’hypothèse première de la grammaire pour les raisons suivantes :
– il existe des langues actuellement parlées qui n’ont aucune grammaire hiérarchique (Jackendoff, 2012 ; Futrell, 2016) ;
– il existe beaucoup de preuves que les symboles ont évolué bien avant la grammaire dans l’histoire linguistique de l’homme ;
– les grammaires hiérarchiques sont dérivées des processus indépendants de la hiérarchie dans l’organisation et la récupération des informations en dehors des langues humaines ;
– les créatures non-humaines semblent utiliser la syntaxe, qui n’est donc pas réservée aux humains ;
– les humains ont évolué de la rigidité cognitive à la flexibilité cognitive et à l’apprentissage basé sur des restrictions culturelles locales et même environnementales.
Au total, les tenants du rôle prédominant de la culture dans l’acquisition du langage ont tendance à penser que le langage n’est pas un organe du cerveau mais un outil culturel (Everett, 2017) évoluant dans le temps et modifiant le moins possible les traits biologiques de ses utilisateurs dans son processus d’évolution.
6. QUELQUES PENSÉES POUR CONCLURE
Nous prenons en compte tous les aspects et nous sommes fermement convaincus que la linguistique évolutive et son caractère interdisciplinaire mèneront la recherche dans le domaine de l’acquisition du langage dans les années à venir. Néanmoins, notre propre perspective sur l’acquisition du langage est formée par l’approche biolinguistique (Berwick & Chomsky, 2017). Le langage est fondamentalement un système biologique et devrait être étudié à l’aide des méthodologies des sciences naturelles. Bien qu’on soit encore loin de comprendre pleinement la base biologique du langage, nous soutenons que le développement du langage, comme tout système biologique, est formé à la fois par l’expérience particulière sur la langue acquise de l’environnement externe et par les restrictions internes à toutes les structures linguistiques. Étant donné l’histoire extrêmement courte de H. sapiens et l’apparition subséquente d’une capacité computationnelle spécifique à l’espèce liée au langage, l’évolution du langage doit avoir été fondée sur d’autres systèmes cognitifs et perceptuels partagés entre espèces et domaines cognitifs. Même si la capacité chez l’homme d’acquérir un langage sophistiqué est unique dans le règne animal et malgré la discontinuité dans les capacités de communication entre les humains et les autres primates, le langage n’est construit que sur des bases génétiques anciennes, qui sont éclairées par la génomique comparative (et bien évidemment enrichies par l’impact socioculturel de l’évolution humaine). L’architecture génétique de la faculté du langage est également explorée par des recherches sur les troubles neurodéveloppementaux qui perturbent le processus naturel de l’acquisition du langage.
Dans cet article, on a parlé des théories les plus importantes sur l’apparition des langues et des derniers développements sur la base génétique de l’acquisition du langage en essayant d’apporter la preuve de la perspective interdisciplinaire que devrait avoir la recherche moderne sur l’acquisition du langage. Il est clairement nécessaire d’identifier d’autres gènes et d’utiliser davantage les systèmes modèles pour mieux comprendre les circuits neuronaux de la communication vocale et leur liaison avec les fondements culturels du langage. Cela sera plus efficacement réalisé dans le champ de la linguistique évolutive avec la coopération des linguistes théoriques qui, en proposant et en améliorant leurs théories de l’acquisition du langage, forment la liaison entre les disciples entourant la recherche sur l’émergence et le développement du langage.
L’utilisation de patients humains ayant des lésions dans les zones cérébrales liées à la parole et l’utilisation accrue d’approches non invasives comme le MEG et le fMRI dans le but d’étudier la parole auprès des populations neurotypiques pourraient aider exceptionnellement dans cette direction. Pourtant, des corrélations entre le fMRI au repos et l’expression des gènes ont récemment été découvertes (Hawrylycz, 2015 ; Richiardi, 2015 ;Wang, 2015), des études à venir qui combinent de telles approches avec l’imagerie de la parole, y compris les enquêtes qui vont au-delà des domaines traditionnels de la parole et du langage (Blank, 2016), pourraient fournir un aperçu des corrélats génomiques de la parole et une compréhension plus profonde des réseaux cérébraux liés à la parole. Enfin, nous croyons que l’utilisation de modèles animaux et l’étude des circuits vocaux chez les primates non humains seront essentielles pour le progrès dans ce domaine.
Nous ne proposons pas de traiter le langage comme un système abstrait séparé de l’expérience humaine. Au contraire, nous rejoignons Pagel (2009) quand il commente le parallélisme de l’évolution biologique avec l’évolution du langage Tableau 1, en observant que le langage ne fonctionne pas comme un système équipé de mécanismes de correction d’erreur intégrés et qu’il est, par conséquent, susceptible d’effets multidimensionnels d’emprunts et d’autres influences qui pourraient corrompre son signal. Mais, comme c’est le cas pour le génome aussi, les langues qu’on utilise aujourd’hui, sont les vainqueurs d’un processus long et dur d’expérimentation de la part de leurs locuteurs. Et nous n’avons aucun raison de ne pas croire que leur évolution continuera au fil des besoins sociaux et génético-anatomiques de leurs utilisateurs, qui continuent – perpétuellement – leur lutte pour la survie physique.
Notes
[1] Le million d’années, en abrégé Ma, est une unité de mesure du temps , utilisée dans diverses disciplines des sciences de la Terre et de l’Univers pour quantifier des durées géologiques ou astronomiques. Un parcours du temps géologique est proposé chez Guy (2002). En ligne : https://hal.archives-ouvertes.fr/hal-00530143/file/le_temps_geologique_bguy.pdf
[2] ka (pour kiloannus) : unité de temps égale à mille, ou 103 années.
[3] Études GWAS : Les études d’association génétique pangénomiques (genomewide association study, GWAS) (Debette, 2012 ; NIH, 2015) ont pour objectif d’identifier des facteurs de susceptibilité génétiques des maladies multifactorielles. Elles consistent à comparer la fréquence de centaines de milliers de variants génétiques distribués sur l’ensemble des chromosomes entre un groupe de cas atteints de la maladie et un groupe de témoins, en utilisant des technologies de génotypage à haut débit. Il s’agit d’une approche « agnostique », sans hypothèse préalable sur les gènes d’intérêt contrairement aux études d’association génétique de type gène candidat. Le nombre très important de tests statistiques effectués nécessite une puissance de calcul importante et une correction sur tests multiples, une valeur p < 5 × 10-8 étant généralement considérée comme significative.
Références bibliographiques
Ackermann, H., Hage, S. R., Ziegler, W. (2014). Brain mechanisms of acoustic communication in humans and nonhuman primates : an evolutionary perspective. Behav. Brain Sci., 37, 529–546
Armeni, K., Willems, R., Frank, S. (2017). Probablistic language models in cognitive neuroscience : Promises and pitfalls. Neuroscience & Behavioral Reviews, 83, 579-588
Bar-Yosef, O. (2017). Can archaeology tell us about the evolution of cognition and language ? Jouurnal of Neurolinguistics, 43(B), 22-227
Bearden, C.E.& Thompson, P.M. (2017). Emerging global initiatives in neurogenetics : The Enhancing Neuroimaging Genetics through Meta-analysis (ENIGMA) Consortium. Neuron, 94, 232–236
Berwick, R., Chomsky, N. (2017). Why only us : Recent questions and answers. Journal of Neurolinguistics, 43, 166-177
Berwick R.C., Friederici A.D., Chomsky N., Bolhuis J. (2013a). Evolution, brain, and the nature of language. Trends in Cognitive Sciences, 17(2), 89-98
Berwick, R. C., Hauser, M. D. & Tattersall, I. (2013b). Neanderthal language ? Just-so stories take center stage. Front. Psychol. 4, 671-672
Blank, I., Balewskia, Z., Mahowalda, K., Fedorenko, E. (2016). Syntactic processing is distributed across the language system. Neuroimage, 127, 307–323Boris S. (2017).The Primary Language of the Human Brain. Procedia Computer Science, 111, 448-462
Button, K.S., Ioannidis, J., Moktysz, C., Nosek, B et al. (2013). Power failure : why small sample size undermines the reliability of neuroscience. Nat. Rev. Neurosci., 14, 365–376
Carrion-Castillo, A., Franke, B. & Fisher, S. E.(2013). Molecular genetics of dyslexia : an overview. Dyslexia, 19, 214–240
Chabout, J., Sarkar, A., Patel, S.R., Radden, T. et al. (2016). A Foxp2 mutation implicated in human speech deficits alters sequencing of ultrasonic vocalizations in adult male mice. Front. Behav. Neurosci., 10, 197-215
Chen, Y.C., Chen, YC., Kuo, HY., Bornschein, U. et al. (2016). Foxp2 controls synaptic wiring of corticostriatal circuits and vocal communication by opposing Mef2c. Nat. Neurosci., 19, 1513–1522
Chomsky, N. (2006). Language and Mind. Cambridge : Cambridge University Press
Chomsky, N. (1986). Knowledge of Language : Its Nature, Origin and Use. New York : New York
Chomsky, N. (1959). A Review of B.F. Skinner’s Verbal Behaviour. Language, 48-63
Christiansen, M.H., Chater, N. (2008). Language as shaped by the brain. Behavioral and Brain Sciences, 31(5), 489–509
Christiansen, M.H., Kirby, S. (2003). Language evolution : consensus and controversies. Trends in Cognitive Sciences, 7(7), 300–307
Clark, E.V.(2003). First Language Acquisition. Cambridge : Cambridge University Press
Cook, V.J., Newson, M. (2007). Chomsky’s Universal Grammar. New Jersey : Blackwell Publishing
Corballis, M. (2017). Language Evolution : A Changing Perspective. Trends in Cognitive Sciences, 21(4), 229-236 (and references herein)
Coupe, C., Hombert, J.-M. (2002). Language at 70,000 BP : evidence from sea-crossings. Paper Presented at the 4th International Conference on the Evolution of Language, Harvard in https://books.google.gr/books?id=luRZu3QPQloC&pg=PA92&lpg=PA92&dq=language+and+sea-crossing&source=bl&ots=OXjCkNjsp8&sig=0SCNPp7NMpB6PRD0TQdIg-RbPtQ&hl=en&sa=X&ved=0ahUKEwiUkICfl_XYAhVDKuwKHd3uBAoQ6AEINDAC#v=onepage&q=Coupe&f=false
Croft, W. (2000). Explaining Language Change : An Evolutionary Approach. London : Longman.
Darwin, C. (1859). The Origin of Species. En ligne : http://darwin-online.org.uk/converted/pdf/1861_OriginNY_F382.pdf
Debette, S. (2012). Comment lire une étude d’association génétique pangénomique (GWAS) ? Sang Thrombose Vaisseaux, 24(5), 240-247
Dediu, D. & Levinson, S.C.(2013). On the antiquity of language : the reinterpretation of Neandertal linguistic capacities and its consequences. Front. Psychol., 4, 397-414
Deriziotis, P., Fisher, S.E.(2017).Speech and Language : Translating the Genome.Trends Genet., 33(9), 642-656
Descartes, R. (1646/1970). Letter to Marquess of Newcastle. In A. Kenny (Ed.), Descartes’ philosophical letters (p. 206). Oxford : Clarendon Press (A. Kenny, Trans.) in Corballis, M. (2017). The evolution of language : Sharing our mental lives. Journal of Neurolinguistics, 43, 120-132
Descartes, R. (1953), Lettre au marquis de Newcastle du 23 novembre 1646, Œuvres, La Pléiade, Éditions Gallimard, p. 1254-1256.
Dunn, M., Greenhill, S., Levinson, S., Gray R. (2011). Evolved structure of language shows lineage-specific trends in word-order universals. Nature, 473, 79–82
Enard, W. (2011) FOXP2 and the role of cortico-basal ganglia circuits in speech and language evolution. Curr. Opin. Neurobiol., 21, 415–424
Enard, W., Pizeworski, M., Fisher, S.E., Lai, C.S. et al (2002). Molecular evolution of FOXP2, a gene involved in speech and language. Nature, 418, 869–872
Estruch, S.B., Graham, S., Deriziotis, P., Fisher, S.. (2016). The language-related transcription factor FOXP2 is post-translationally modified with small ubiquitin-like modifiers. Sci. Rep., 6, 20911
Evans, N., Levinson, S. (2009). The Myth of Language Universals : Language diversity and its importance for cognitive science. Behavioral and Brain Sciences, 1-62
Everett, D.L.(2017). Grammar came later : Triality of patterning and the gradual evolution of language. Journal of Neurolinguistics 43,133-165
Everett, D. L.(2016). Dark Matter of the Mind : The Culturally Articulated Unconscious. Chicago : University of Chicago Press
Everett, D. L.(2012). Language : The Cultural Tool. New York : Pantheon Books
Everett, D. L.(2005). Cultural Constraints on Grammar and Cognition in Pirahã : Another Look at the Design Features of Human Language. Current Anthropology, 46(4), 621–646
Fedorenko, E., Nancy Kanwisher, N. (2009). Neuroimaging of Language : Why Hasn’t a Clearer Picture Emerged ? Language and Linguistics Compass. 3,10.1111/j.1749-818x.2009.00143.x. En ligne :http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.208.4084&rep=rep1&type=pdf
Fisher, S.E.,Scharff, C. (2009). FOXP2 as a molecular window into speech and language. Trends Genet. 25, 166–177
Fisher, S.E.,Marcus, G.F.(2006). The eloquent ape : genes, brains and the evolution of language. Nat. Rev. Genet., 7, 9–20
Fitch, W.T.(2010). The Evolution of Language. Cambridge : Cambridge University Press.
French, C.A., Fisher, S.E.(2014). What can mice tell us about Foxp2 function ? Curr. Opin. Neurobiol., 28, 72–79
French, C.A., French C.A., Jin, X., Campbell, T.G. et al. (2012). An aetiological Foxp2 mutation causes aberrant striatal activity and alters plasticity during skill learning. Mol. Psychiatry, 17, 1077–1085
Fuentes, A. (2016). The extended evolutionary synthesis, ethnography, and the human Niche : Toward an integrated anthropology. Current Anthropology, 57(Suppl. 13), S13-S26
Futrell, R., Stearns, L., Piantadosi, S. T., Everett, D. L., Gibson, E. (2016). A corpus investigation of syntactic embedding in Pirahae. PLoS One, 11(3) : e0145289
Gialluisi, A., Guadalupe, T., Francks, C., Fisher, SE. (2017). Neuroimaging genetic analyses of novel candidate genes associated with reading and language. Brain Lang., 172, 9-15
Gong, T., Shuai, L., Comrie, B. (2014). Evolutionary linguistics : theory of language in an interdisciplinary space. Language Sciences, 41 ; 243–253
Gould, S.J., Vrba, E.S.(1982). Exaptation : a missing term in the science of form. Paleobiology, 8(1), 4–15
Graham, S.A.& Fisher, S.E.(2015). Understanding language from a genomic perspective. Annu. Rev. Genet., 49, 131–160
Graham, S. A.& Fisher, S. E.(2013). Decoding the genetics of speech and language. Curr. Opin. Neurobiol., 23, 43–51
Greenberg, J.H.(1963). Some universals of grammar with particular reference to the order of meaningful elements. In : Greenberg, J.H.(Ed.), Universals of Language (pp. 73–113). Cambridge, MA : MIT Press
Groszer, M., Keays, D.A., Deacon R.M.J., de Bono, J.P. et al. (2008). Impaired synaptic plasticity and motor learning in mice with a point mutation implicated in human speech deficits. Curr. Biol., 18, 354–362
Hagoort P. (2017). The core and beyond in the language-ready brain. Neuroscience & Behavioral Reviews, 81B, 194-204
Hauser, M.D., Barner, D., O’Donnell, T. (2007). Evolutionary linguistics : a new look at an old landscape. Language. Learning and Development, 3(2), 101–132
Hauser, M.D., Chomsky, N., Fitch,W.T.(2002). The Faculty of Language : What Is It, Who Has It, and How Did It Evolve ? Science, 298, 5598, 1569 – 1579
Hawrylycz, M., Miller, J.A., Menon, V., Feng, D. et al. (2015). Canonical genetic signatures of the adult human brain. Nat Neurosci. 2015 ; 18, 1832–1844
Hibar, D.P. et al. (2015). Common genetic variants influence human subcortical brain structures. Nature, 520, 224–229
Hockett, C.F.(1960). The origin of speech. Scientific American, 203, 88–96
Hoogman, M., Guadalupe, T., Zwiers, M.P., Klarenbeek, P. et al. (2014). Assessing the effects of common variation in the FOXP2 gene on human brain structure. Frontiers in Human Neuroscience, Article 473. En ligne : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4076884/pdf/fnhum-08-00473.pdf
Hulme, C., Snowling, M. J. (2013) The interface between spoken and written language : developmental disorders. Philos. Trans. R. Soc. Lond. B. Biol. Sci., 369, 20120395 ; doi : 10.1098/rstb.2012.0395
Hurford, J.R.(2012). The Origins of Grammar. Oxford : Oxford University Press, Oxford.
Jackendoff, R., Wittenberg, E. (2012). Even simpler syntax. En ligne :https://depts.washington.edu/lingconf/abstracts/JackendoffandWittenberg.pdf.
Janik, V.M. (2014). Cetacean vocal learning and communication. Curr. Opin. Neurobiol., 28, 60-65
Jiménez-Bravo, M., Marrero, V., Benitez-Burraco, A. (2017).An oscillopathic approach to developmental dyslexia : From genes to speech processing. Behavioural Brain Research, 329, 84-95
Johansson, S. (2014). Neanderthals did speak, but FOXP2 doesn’t prove it. Behav. Brain Sci., 37, 577–604
Ke, J., Holland, J.H.(2006). Language origin from an emergentist perspective. Applied Linguistics, 27(4), 691–716
Kirby, S. (2007). The evolution of language. In : Dunbar, R., Barrett, L. (Eds.), The Oxford Handbook of Evolutionary Psychology. Oxford : Oxford University Press
Klein, R.G.(2017). Language and human evolution. Journal of Neurolinguistics, 43, 204-221
Konopka, G., Bomar, JM, Coppola G., Jonsson, Z.O. et al. (2009) .Human-specific transcriptional regulation of CNS development genes by FOXP2. Nature, 462, 213–217
Krause, J., Lalueza, C., Ludovic, O., Enard, W. et al. (2007). The derived FOXP2 variant of modern humans was shared with Neandertals. Curr. Biol., 17, 1908–1912
Lai, C. S., Fisher, S. E., Hurst, J. A., Vargha-Khadem, F. (2001). A forkhead-domain gene is mutated in a severe speech and language disorder. Nature, 413, 519–523
Le Fevre, A.K., Taylor, S., Malek, NH., Horn, D. et al. (2013). FOXP1 mutations cause intellectual disability and a recognizable phenotype. Am. J. Med. Genet., A 161A, 3166–3175
Li, S. et al. (2004). Transcriptional and DNA binding activity of the Foxp1/2/4 family is modulated by heterotypic and homotypic protein interactions. Mol Cell Biol., 24, 809–822
Lieberman P. (2006). Toward an Evolutionary Biology of Language. Cambridge, MA : Belknap Press of Harvard University Press
Lozano, R., Vino, A., Lozano, C., Fisher, ES., Deriziotis, P. (2015). A de novo FOXP1 variant in a patient with autism, intellectual disability and severe speech and language impairment. Eur. J. Hum. Genet. 23, 1702–1707
Luuk, E.,Luuk, H. (2014). The evolution of syntax : Signs, concatenation and embedding. Cognitive Systems Research., 27, 1-10
Mars, R., Eichert, N., Jbabdi, S., Verhagen, L. (2018). Connectivity and the search for speacializations in the language-capable brain. CurrOpinBehavSci, 21, 10-26
Mattingly, I. G.(1972). Reading, the linguistic process and linguistic awareness. In Kavanagh J. F., Mattingly I. G.(Eds.), Language by ear and by eyes : the relationships between speech and reading (pp. 133–147). Cambridge, MA : MIT Press
Meyer, M., Kircher, M., Gansauge, M.-T., Li, H. et al. (2012). A high-coverage genome sequence from an archaic Denisovan individual. Science, 338, 222-226
Moreno, A., Limousin, F., Dehaene, S., Pallier, C. (2018). Brain correlates of constituent structure in sign language comprehension. Neuroimage, 167,151-161
Morgan, A., Fisher, S.,Scheffer, I., Hildebrand, M. (2016). FOXP2-related speech and language disorders. In Pagon, R.A.(Eds.), Gene Reviews (pp. 1993–2017), Seattle, WA : University of Washington
Mozzi A., Forni D., Clerici M., Pozzoli U. et al (2016). The evolutionary history of genes involved in spoken and written language : beyond FOXP2. Scientific Reports, 6, Article number : 22157, doi:10.1038/srep22157
Mufwene, S.S.(2008). What do creoles and pidgins tell us about the evolution of languages ? In : Laks, B. (Ed.), Origin and Evolution of Languages : Approaches, Models, Paradigms (pp 272-297). London : Equinox. En ligne : http://mufwene.uchicago.edu/publications/CREOLES-LGEVOLUTION-Revisions-1.pdf
Mukamel, Z., Konopka, G., Osborn, GE.,Donh, H. et al. (2011). Regulation of MET by FOXP2, genes implicated in higher cognitive dysfunction and autism risk. J. Neurosci., 31, 11437–11442
Müller, S. (2013). Unifying everything : Some remarks on simpler syntax, construction grammar, minimalism, and HPSG. Language, 89(4), 920–950
Murugan, M., Harward, S., Scharff, C., Mooney, R. (2013) Diminished FoxP2 levels affect dopaminergic modulation of corticostriatal signaling important to song variability.
Neuron 80, 1464–1476. National Human Genome Research Institute (2015). Genome-Wide Association Studies. En ligne : https://www.genome.gov/20019523/genomewide-association-studies-fact-sheet/
Pagel, M. (2009). Human language as a culturally transmitted replicator. Nature Reviews|Genetics, 10, 405-415
Paracchini, S. (2011). Dissection of genetic associations with language-related traits in population-based cohorts. J. Neurodev. Disord., 3, 365–373
Peterson, R. L.,Pennington, B. F.(2012). Developmental dyslexia. Lancet, 379, 1997–2007
Piantadosi, S., Tily, H., Gibson, E. (2012), The communicative function of ambiguity in language. Cognition, 122,280–291
Pinker, S.,Jackendoff, R. (2005). The Nature of the Language Faculty and its Implications for Evolution of Language : Reply to Fitch, Hauser, and Chomsky. Cognition : International Journal of Cognitive Science, 97(2), 211-225
Pinker, S. (1995). The Language Instinct : The New Science of Language and Mind. Harmondsworth : Penguin
Pinker, S., Bloom, P. (1990). Natural language and natural selection. Behavioral and Brain Sciences 13(4), 707–784
Plaza, M. (2016). Psychopathologie du langage. Enfance et Psychopathologie (10eedition)
Reich, D., Green, R. E., Kircher, M., Krause, J. et al. (2010). Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature, 468(7327), 1053-1060
Richiardi, J., Altmann, A., Milazzo, A.C., Chang, C. et al. (2015). Brain Networks. Correlated gene expression supports synchronous activity in brain networks. Science, 348, 1241–1244
Roll, P., Vernes, SC, Bruneau, N, Cillario J, et al. (2010). Molecular networks implicated in speech-related disorders : FOXP2 regulates the SRPX2/uPAR complex. Hum. Mol. Genet. 19, 4848–4860
Schizophrenia Working Group of the Psychiatric Genomics Consortium (2014). Biological insights from 108 schizophrenia-associated genetic loci. Nature 511, 421–427
Skinner, B.F.(1957). Verbal Behavior. New York : Prentice Hall
Sollis, E., Graham, S., Vino, A., Froelich, H.et al. (2016). Identification and functional characterization of de novo FOXP1 variants provides novel insights into the etiology of neurodevelopmental disorder. Hum. Mol. Genet., 25, 546–557
Steels, L. (2011). Modeling the cultural evolution of language. Physics of Life Reviews, 8, 339–356
Stout D., Chaminade, T. (2012). Stone tools, language and the brain in human evolution. Philos Trans Soc Lond Bio Sci, 367(1585), 75-87
Stout, D. (2011). Stone toolmaking and the evolution of human culture and cognition. Phil. Trans. R. Soc. B, 366, 1050–1059
Stout, D., Chaminade T. (2009). Making tools and making sense : complex, intentional behaviour in human evolution. Camb. Archaeol. J..19, 85–96
Strauss, K.A., Puffenberger, EG, Huentelman, MJ, Gottlieb, S et al. (2006). Recessive symptomatic focal epilepsy and mutant contactin-associated protein-like 2. N. Engl. J. Med. 354, 1370–1377
Tatersall, I. (2017). The material record and the antiquity of language. Neuroscience & Behavioral Reviews, 2017, 81(B), 247-254
Thomason,S., Kaufman, T. (1988). Language Contact, Creolization, and Genetic
Linguistics.California. University of California PressTomasello, M.,Silke B. (2009). Flexibility in the Semantics and Syntax of Children’s Early Verb use. Monographs of the Society for Research in Child Development, 74(2), 113-126
Tomasello, M. (2008). The Usage-based theory of language acquisition. In Bavin, E, (Ed.) The Cambridge handbook of child language (pp.69-88). New York : Cambridge University Press
Tomasello, M. (2003). Constructing a Language : A Usage-Based Theory of Language Acquisition. Cambridge MA : Harvard University Press
Tomasello, M. (2000). Do young children have adult syntactic competence ? Cognition, 74,209-253
van der Lely, H. K., Pinker, S. (2014). The biological basis of language : insight from developmental grammatical impairments. Trends Cogn. Sci., 18, 586–595
Vernes, S.C., Newbury, D., Abrahams, B., Winchster, L. et al. (2008). A functional genetic link between distinct developmental language disorders. N. Engl. J. Med. 359, 2337–2345
Walker, R.M., Hill, A. E., Newman, A. C., Hamilton, G. et al. (2012). The DISC1 promoter : characterization and regulation by FOXP2. Hum. Mol. Genet., 21, 2862–2872
Wang, G.Z., Belgard, T.G., Mao, D., Chen, L. et al. (2015). Correspondence between Resting-State Activity and Brain Gene Expression. Neuron,88, 659–666
White, T.D., Asfaw, B., Degusta, D., Gilbert, H. et al (2003). Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature, 423, 742–747
Wohlgemuth, S., Adam, I., Scharff, C. (2014). FoxP2 in songbirds. Curr. Opin. Neurobiol., 28, 86–93